![]()


|
|
|
COMPLESSITÀ E DOLORE
GIUSEPPE SERAFINI - ALESSANDRO F. SABATO*
Università degli Studi di Roma Tor Vergata
Scuola di Specializzazione in Anestesia e Rianimazione (Dir. Prof. G. Leonardis)
*Cattedra di Anestesia Generale e Speciale Odontostomatologica, Servizio di Fisiopatologia e Terapia del Dolore (Resp. Prof. A.F.Sabato)
Gli impressionanti successi della biologia molecolare nelle ultime decadi hanno portato alcuni autori a ritenere che le sole investigazioni biologiche autenticamente degne dello statuto scientifico siano quelle in grado di spiegare i fenomeni biologici nei termini dei componenti elementari e dei sottesi processi fisico-chimici. Le aspettative di tali autori si spingono fino all’ideale di poter esaustivamente spiegare il comportamento di interi sistemi sulla base di leggi avanzate in grado di chiarire il comportamento degli atomi e delle molecole che lo compongono. Non vogliamo addentrarci, in questo ambito, nelle controversie teoretiche del riduzionismo biologico. Riteniamo che il solo criterio di validità di una strategia di ricerca sia il suo successo. Vale a dire, i metodi analitici così come quelli sintetici, cioè lo studio dei fenomeni viventi ai livelli più bassi o più alti di complessità, sono giustificati se incrementano la nostra comprensione di un fenomeno, che in medicina si traducono sempre in efficaci, o quanto meno più efficaci di quelle a quel momento disponibili, strategie terapeutiche. Al di là di posizioni estreme, riconosciamo la validità di distinti livelli di descrizione scientifica (dai quark all’analisi di società intere), ciascuno dei quali non è logicamente riducibile all’altro: ogni livello di lettura ha proprie leggi, e nessuno può essere ricondotto, al momento, ad una scienza di base che comprenda tutte le altre.
Ci sembra ragionevole, ed inevitabile, prospettare così la rilevanza e la necessità di evidenziare:
Il livello di descrizione scientifica su cui questo lavoro intende collocarsi è quello dei sistemi complessi, volti ad indagare i paesaggi della dinamica "non-lineare", cioè a paesaggi dell’energia più complicati di quelli corrispondenti ai modelli della fisica dell’ordine di Newton-Laplace. Questa dinamica "non-lineare" è alla base dell’apparizione, all’interno di sistemi ad evoluzione temporale deterministica ben definita, del caos: imprevedibili scarti dalla traiettoria (o dal comportamento) ideale, che in tempi lunghi mutano scenari (configurazioni) e aspettative (evoluzione).
Questa non-linearità è anche la base esplicativa dell’auto-organizzazione, fenomeno per cui, nei sistemi biologici, a partire da una situazione disordinata, si evolve spontaneamente verso uno stato più ordinato, attraverso l’organizzazione (schemi), mediante meccanismi di feedback (omeostasi), di diversi sotto-sistemi interagenti a più livelli.
Il quadro concettuale, non consueto, in cui svolgeremo le nostre riflessioni è quello dello studio delle leggi della complessità in quanto tale, non solo i sistemi complessi nella loro specifica composizione.
"Non so definirla, ma se la vedo la riconosco". Parole usate per la pornografia, ma che ben si adattano alla complessità, motivo di frontiera della dinamica non-lineare.
Quello che intuitivamente sembra ben sviluppato al fine di individuare un sistema complesso, alla luce dei tentativi di formalizzarlo, si trasforma in un vago senso di incapacità nell’afferrare gli aspetti essenziali del problema. Certamente il termine complessità è utile, più di ogni altro, per descrivere i sistemi altamente organizzati. E volendo sviluppare uno schema speculativo è necessario considerare un insieme di concetti teoretici strettamente collegati alla nozione di complessità (gerarchie, adattamento, biforcazioni, autorganizzazione, ecc.) per i quali rimandiamo ai nostri lavori precedenti (31.Sabato e Serafini, 1996; 32.Serafini e Sabato, 1996; 33.Serafini e Sabato, 1997; 34.Serafini, 1997), che fanno emergere la complessità non come proprietà intrinseca di un sistema isolato (chiuso), ma come proprietà latente o implicita di un sistema, che si rende manifesta solo attraverso l’interazione tra il sistema dato con un altro. In questo senso, è probabilmente più appropriato considerare la complessità più come proprietà dell’interazione che del sistema, sebbene sia chiaramente associata ad entrambi. Al di là di dettagliate considerazioni sulla complessità nei fenomeni naturali e umani, è utile considerare perché una comprensione più profonda della complessità, in sé, sia di importanza e teorica e pratica. La ragione basilare è inerente alla necessità di semplificare al fine di comprendere e dirigere (controllare) un fenomeno. E dal momento che la maggior parte delle nostre conoscenze, e virtualmente tutti i processi di controllo sono basati su un modello (mentale, matematico, fisico, o altro) del sistema (fenomeno) in studio, la semplificazione traduce questo desiderio di ottenere una rappresentazione equivalente, ma ridotta, del modello originale del sistema. Questo procedimento può implicare che alcune variabili originali siano omesse, che altre siano invece aggregate, che interazioni deboli siano ignorate, che vengano considerate costanti lente variazioni, ed un insieme di altri sotterfugi. Tutte queste tecniche di semplificazione sono orientate a ridurre i gradi di libertà1; che il sistema ha a disposizione per interagire con il suo ambiente. Una utile teoria della complessità dei sistemi dovrebbe farci acquisire conoscenza ponendo dei limiti ai processi di riduzione, o meglio, fornendo delle condizioni in base alle quali un modello anche se ridotto possa funzionare e forse anche suggerire nuove, più semplici rappresentazioni che possano essere valide quando la semplificazione "naturale" fallisse.
Nota
1 Numero dei parametri indipendenti necessari a specificare la configurazione di un sistema.
Chiarito, o almeno premesso ciò, possiamo chiederci: già, ma cos’è un comportamento complesso? O meglio, in base a quale(i) criteri(o) possiamo riferirci ad un sistema deducendone o meno la sua complessità? Quali sono, cioè, le caratteristiche comportamentali e strutturali distintive di questi sistemi che intuitivamente riteniamo essere complessi? Forse il modo più semplice di affrontare tali questioni è quello di cominciare dal loro opposto: quali caratteristiche associamo a sistemi semplici?
Le proprietà più evidenti di un sistema semplice sono:
Un comportamento predicibile. È inutile aspettarsi sorprese, nessun cambiamento auto-indotto è possibile. Data la costanza e l’ergodicità di un sistema che conserva energia, inoltre, è sempre possibile calcolare la probabilità che il sistema è in qualche specifico volume delle fasi2 corrispondente a qualche stato macroscopico (17.Kauffman, 1993; 18.Kauffman, 1995). È, cioè, deterministicamente prevedibile. Un sistema semplice esibisce, quindi, un modello di comportamento che è facile dedurre conoscendo le forze esterne che agiscono sul sistema.
Al contrario, i sistemi "non-ergodici"3;, complessi, manifestano gradi crescenti di complessità (36.Soodak and Iberall, 1987; 4.Casti and Karquist, 1985). Esibiscono comportamenti contro-intuitivi, apparentemente acausali e pieni di sorprese impredicibili. Se rivolgiamo, infatti, l’attenzione ad un sistema biologico, non possiamo che osservare la sua evolutività dinamica. Vale a dire, non solo come sistema termodinamicamente aperto e dissipativo (ossia che dissipa energia in calore) diviene sempre più complesso, ma, qualunque sia il livello di complessità raggiunto, il processo dell’informazione al suo interno porta all’emergenza di proprietà qualitativamente nuove (proprietà emergenti), a seconda del grado di complessità, che fa esprimere il sistema in modelli di comportamento nuovi e progressivamente imprevedibili, senza che comunque esso possa vagare a caso ed ergodicamente nello spazio delle sue possibilità. Va tenuto presente, infatti, come già scritto, che un sistema biologico complesso dissipativo (dal sistema genomico ai sistemi d’organo, al sistema immunitario, alle reti neurali, agli ecosistemi) ha sempre attrattori4(o, come vedremo successivamente, schemi).
Note
2 (O spazio delle fasi o spazio degli stati).Rappresentazione grafica N-dimensionale in cui ad ogni asse (dimensione) č associata una variabile dinamica (N). Un punto in questo spazio indica lo stato del sistema in un determinato istante. Ogni cambiamento dello stato del sistema produce uno spostamento del punto, che descrive una certa traiettoria.
3 "Non-ergodico" č un termine tecnico per indicare un sistema che ammette molti possibili stati di equilibrio, sistemi cioč in cui esistono obiettivi da raggiungere in mutuo conflitto per cui, se se ne raggiunge uno, si č dovuto rinunciare agli altri. Un esempio tipico č dato dal sistema immunitario: se si orienta, ad es., contro il virus influenzale, si rimane scoperti verso altri attacchi. Altro esempio, a noi piů vicino, č il sistema Nondolore-Dolore (Nd/D), in cui la percezione del sintomo č ordinata gerarchicamente, nel senso che un nuovo o maggiore dolore "scaccia" il precedente.
4 Insieme di punti o stati nello spazio delle fasi verso cui le traiettorie all'interno di un qualche volume dello spazio delle fasi convergono asintoticamente nel tempo.
Ridotto numero di interazioni e circuiti lineari di feedback/feedforward. I sistemi semplici generalmente coinvolgono un piccolo numero di componenti, con auto-interazioni che prevalgono sulle mutue interazioni delle variabili. Possiedono, inoltre, sistemi di regolazione a controllo in feedback (retroazione)/feedforward (informazione anticipata) che sono lineari, in cui cioè si assume che tutte le componenti dinamiche del circuito di controllo sono invarianti nel tempo, con pochi inputs e scarsi disturbi ambientali. I sistemi complessi sono tipicamente composti da un alto numero di variabili fortemente interagenti, e raggruppate in sottosistemi anch’essi interagenti, anche se a minor grado rispetto alle interazioni intra-sottosistema. In essi il comportamento dinamico è sottoposto a processi di regolazione non-lineari, con molti inputs, ed in cui le varie componenti dei circuiti, multipli peraltro, modificano nel tempo il valore dei loro parametri e la loro struttura, al fine di rendere il sistema capace di ristrutturare, o almeno di modificare i modelli di interazione delle sue variabili attraverso un più ampio range di potenziali modelli di comportamento (2.Aida et al., 1985). Vale la pena di sottolineare che nei sistemi viventi l’omeostasi è una particolare forma di regolazione a controllo retroattivo.
In linea molto generale, l’omeostasi rappresenta la capacità dell’organismo nel suo insieme o di sue sub-componenti di conservare costanti, o meglio variabili entro determinati limiti, dei parametri biologici o delle funzioni in modo che tali parametri e funzioni concorrano al buon funzionamento dell’organismo nel suo insieme. In questa ottica le oscillazioni dei parametri soggetti a controllo omeostatico non vanno considerate dei disturbi legati a fluttuazioni casuali e disordinate, inevitabile risultato dello squilibrio tra sistemi di controllo; piuttosto la modulazione di frequenza che esse producono realizza un meccanismo di mutua informazione cellulare, in grado di trasferire le più complesse computazioni, e di contribuire alla sincronizzazione e al coordinamento di grandi gruppi di cellule svolgenti la stessa funzione (24.Meyer, 1991), come tra l’altro è stato osservato nelle oscillazioni dell’attività corticale cerebrale (9.Engel et al., 1992).
Man mano che la complessità dei sistemi deputati a tali scopi si è andata rendendo più evidente con il progresso delle scienze biomediche, il concetto di omeostasi si è ampliato, nel senso che mentre inizialmente la sua applicazione poteva essere delimitata nel campo della fisiologia classica di organo, apparato o sistema (ad es.: circolazione, sistema endocrino, apparato escretore), oggi si può constatare che esiste un’omeostasi anche ad altri livelli, sia sul piano cellulare e molecolare (ad es.: concentrazioni di ioni nel citoplasma, velocità di catalisi di un enzima, ecc.) che sul piano dei sistemi e delle funzioni che controllano l’integrità e la qualità dell’informazione biologicamente significativa (ad es.: sistema immunitario, neurobiologia, sistema nondolore-dolore -Nd/d).
È opportuno ribadire e chiarire il fatto che il concetto di omeostasi non deve essere confuso con equilibrio stazionario. I vari sistemi biologici non sono mai in "equilibrio"; piuttosto, essi vanno soggetti a continue oscillazioni nell’intensità dei fenomeni ad essi correlati, in quanto sono mantenuti lontano dall’equilibrio da un continuo flusso di energia. I sistemi biologici sono oggi visti come tipici sistemi dinamici "dissipativi", nel senso che il loro stato stazionario (steady-state) è mantenuto dal continuo consumo di energia che mantiene l’ordine in uno spazio-tempo limitato, a spese dell’aumento di entropia dell’ambiente circostante (11.Guidotti, 1990; 26.Nicolis e Prigogine, 1991). In altri termini, l’organizzazione biologica genera ed usa strutture spazio-temporali in cui non c’è omogeneità né fase statica ad alcun livello. Lo "stato" stazionario non è uno stato, ma un conglomerato di processi organizzati spazio-temporalmente.
Processo decisionale accentrato. Nei sistemi semplici il controllo, inteso come funzione decisionale, è ristretto ad uno, o comunque pochissimi centri decisionali, l’effetto delle cui azioni sul sistema stesso è solitamente facile da prevedere. In contrasto, il controllo di sistemi complessi tende ad essere molto decentrato in quella che in cibernetica viene definita intelligenza distribuita: se il sistema deve presentare un comportamento coerente, questo avrà origine dalla competizione e dalla cooperazione fra tutte le componenti.
Scomponibilità. Tipicamente un sistema semplice implica scarse interazioni tra i suoi componenti costitutivi, tale che il comportamento del sistema nel suo insieme poco o nulla risente della riduzione di tali interazioni. Può quindi essere scomposto e studiato nei suoi processi indipendenti (o scarsamente interagenti) senza alterare l’integrità fondamentale del sistema. Dall’altra parte invece, un processo complesso è quasi-scomponibile. Supponiamo di avere un sistema complesso di N variabili. Per prevedere il suo comportamento dobbiamo risolvere N equazioni simultaneamente. Supponiamo, comunque, che il sistema sia composto da m sottosistemi stabili ciascuno contenente k variabili, dove mk=N. Laddove il sistema è strutturato in gerarchia (che è l’architettura dominante tra i sistemi naturali), vale a dire che le variabili in ogni particolare sottosistema hanno, mediamente, interazioni relativamente più forti con le altre variabili dello stesso sottosistema, e, mediamente, molto più deboli con le variabili appartenenti agli altri sottosistemi componenti, allora si potrà predire il comportamento di ciascun sottosistema risolvendo solamente le k equazioni simultanee. E avendo trovato le soluzioni per ciascun sottosistema possiamo ottenere la risposta in bassa frequenza (cioè nei tempi brevi , ad alto grado di modularizzazione e a bassa frequenza di inputs) di tutto il sistema risolvendo solamente le m equazioni simultanee, una per ogni sottosistema. In tali condizioni, quindi, più deboli saranno le interazioni tra i sottosistemi, più sarà breve il tempo richiesto per ripristinare l’equilibrio, una volta alterato, all’interno del singolo sottosistema, date però le integrità strutturali in e tra i vari sottosistemi componenti. Ogni alterazione delle connessioni informative tra le variabili o i sottosistemi che lo compongono può alterare stabilmente gli aspetti essenziali e l’armonizzazione del comportamento globale del sistema, soprattutto nelle sue risposte ad alta frequenza (cioè ad alte risoluzioni temporali, con sottosistemi fortemente interagenti e ad alta frequenza di inputs) e adattative. Nei sistemi biologici complessi la patologia può iniziare proprio come modifica di connettività tra gli elementi del sistema globale. Tale mutazione di connessioni può aumentare il disordine del sistema, perché alcuni elementi sfuggono al gioco dei controlli crociati ed iniziano a presentare dinamiche proprie, autonome, soggette più facilmente ad ampie oscillazioni.
Se si modifica il coordinamento del sistema nel suo insieme e con il resto dell’organismo, alcune sub-componenti possono oscillare in modo eccessivo, imprevedibile, generando quindi disordini localizzati, che però possono essere amplificati e trasmessi al resto del sistema o ad altri sistemi in modo disordinato e afinalistico. Al limite, variazioni troppo rapide ed intense delle variabili implicate in un sistema omeostatico possono configurare una situazione di feed-back positivo amplificato fino all’autodistruzione del sistema.
In una rete in cui molti sotto-sistemi omeostatici oscillanti sono interconnessi, l’informazione del sistema intero percorre dei cicli che hanno forme spazio-temporali variabili, fluttuanti, ma sempre riconducibili ad uno schema armonizzato di autorganizzazione globale, con il minor dispendio di energia possibile. Se uno o più elementi di tali reti modificano le connessioni informative, cioè il sistema omeostatico si spezza, o si spezza il flusso d’informazione tra diversi sotto-sistemi, si ha un processo patologico proprio in quanto il sistema modifica afinalisticamente l’armonia e "l’identità" del suo comportamento globale.
Vi sono molti modi con cui un sistema integrato muta connettività e qui ne elenchiamo alcuni a titolo esemplificativo:
Comportamento predicibile, ridotto numero di interazioni e circuiti lineari di feedback/feedforward, processo decisionale accentrato, scomponibilità, caratteristiche di un sistema semplice che, di converso, fanno emergere un quadro in cui la nozione di fenomeno complesso viene ad essere caratterizzato da modelli di comportamento contro-intuitivi indeducibili dalla conoscenza degli inputs ambientali; da un alto numero di variabili fortemente interagenti in una ricca rete di connessioni in feedback/feedforward non-lineari; da processi decisionali decentrati e da un alto livello di modularità. Per caratterizzare compiutamente un sistema complesso a queste caratteristiche occorre aggiungere:
Capacità di adattamento. La capacità cioè del sistema di monitorizzare il suo ambiente e di riconfigurarsi sulla base delle sue osservazioni al fine di rendere più efficaci le sue funzioni.
Organizzazione gerarchizzata. Il sistema tende ad essere strutturalmente organizzato in maniera stratificata in modo tale che l’informazione e l’attività del livello inferiore (peraltro più semplice e più piccolo del suo prodotto) risultano inputs per i livelli superiori, mentre la direzione ed il controllo passano dai livelli più alti a quelli più bassi.
Biforcazioni5 e proprietà emergenti. Il sistema complesso esibisce, a seconda del grado di complessità raggiunto, modelli di comportamento e di organizzazione strutturale e funzionale del tutto nuove e rispetto a pattern precedenti, e rispetto alle proprietà singolarmente possedute dai suoi componenti. Tali qualità emergenti rappresentano punti di biforcazione dove le precedenti descrizioni del sistema risultano inefficaci, e nuove leggi, nuovi concetti e nuove generalizzazioni, non riducibili alle precedenti, sono necessarie
Nota
5 Biforcazioni nel comportamento di un sistema dinamico sono improvvisi e vistosi cambiamenti di traiettoria ed attrattori, senza stati transitori intermedi, che fanno seguito a piccole modificazioni nei parametri di funzionamento del sistema stesso. Un esempio intuitivo di cosa significhi una biforcazione ci č fornito dalla fluidodinamica: un flusso puň essere laminare per un ampio intervallo di velocitŕ del liquido, ma al crescere della velocitŕ si arriverŕ ad un punto in cui insorge la turbolenza (punto di biforcazione).
Possiamo affermare che il nucleo del problema, o meglio, l’osservazione che ha fatto sì che il sistema delle conoscenze acquisite e dell’interpretazione dei fenomeni naturali venissero sconvolti, è quella per cui in un sistema complesso, ad evoluzione temporale "deterministica" ben definita (sistemi, cioè, dinamici), l’interazione delle sue sub-componenti su una scala di osservazione, porta ad un comportamento globale complesso su una scala più ampia, che in generale non può essere previsto dalla conoscenza delle singole componenti. Non solo, ma che tale imprevedibilità non è dovuta alla mancanza di informazioni sul sistema stesso, non può essere colmata dall’acquisizione di nuove informazioni, bensì è una proprietà intrinseca dei sistemi fisici, che si manifesta in modo più o meno evidente, con comportamenti e proprietà nuove, a seconda delle condizioni interne o esterne al sistema stesso, in ultima analisi del suo livello di organizzazione o meglio di complessità.
In generale, l’individuazione di gradi differenti di complessità risponde, più che ad un’esigenza tassonomica (pur sempre di estrema utilità dialogica), all’evidenza di una dinamica non-lineare appartenente a sistemi con molteplici stati di equilibrio, in cui possono coesistere molte configurazioni diverse non riducibili al gioco dei componenti.
Occorre così operare, almeno nell’ambito dei sistemi biologici, una distinzione tra complessità subcritica ( od ordinata), critica (o propriamente complessa) e caotica.
Nella complessità subcritica il sistema confina spontaneamente i suoi comportamenti dinamici in un volume relativamente piccolo all’interno del suo spazio delle fasi. Vale a dire assume comportamenti deterministici, prevedibili nel tempo e definibili da equazioni strettamente deterministiche, in cui la maggior parte dei cambiamenti nei parametri di funzionamento del sistema (biforcazioni) non modifica drammaticamente il suo comportamento dinamico, ma lo lascia all’interno di un piccolo volume del suo spazio delle possibilità. Ha quindi la proprietà di essere strutturalmente stabile: piccoli cambiamenti nello spazio dei parametri provoca, per lo più, piccole modificazioni nel comportamento dinamico; e quella di essere strutturalmente o funzionalmente modulare: i sottosistemi da cui è composto hanno un basso grado di connessione, non sono cioè fortemente interagenti ma relativamente indipendenti. Questo vuol dire anche che un sistema in fase ordinata, laddove "perturbato", assume un comportamento non ergodico, all’interno di volumi ben confinati nello spazio delle possibilità, tendendo fortemente a ritornare allo stesso comportamento iniziale e a mantenerlo una volta finita la perturbazione: è quindi omeostaticamente stabile. Nel linguaggio dei sistemi dinamici il comportamento di un sistema complesso in questa fase va a cadere in un attrattore punto6 o in attrattori periodici7 stabili. Così come corrisponde, volendo proseguire le analogie, alle classi I e II di automi cellulari (39.Wolfram, 1983; 40.Wolfram, 1984; 20.Langton, 1986; 22.Langton, 1992), e, in fisica, al regime solido.
Note
6 Singolo stato, che non cambia nel tempo, verso cui convergono traiettorie differenti.
7 O ciclo limite: insieme di stati all'interno di cui il sistema oscilla ripetitivamente.
Nella fase di complessità critica il sistema si muove in una regione di comportamenti complessi speciali che separa l’ordine dal caos. Un luogo di confine dove il comportamento ordinato è continuamente scalzato da comportamenti straordinariamente complessi, senza mai dissolversi nella turbolenza caotica. Il cosiddetto "margine del caos" (20.Langton, 1986; 21.Langton, 1990; 22.Langton, 1992; 27.Packard, 1988; 17.Kauffman, 1993), dove sembra che un sistema dinamico dissipativo assuma i comportamenti più flessibili, più complessi, più integrati e compia i passi evolutivi più importanti per sé e per i sistemi ad esso correlati. E’ una fase in cui il sistema è sufficientemente ordinato da esibire una qualche stabilità strutturale,- è cioè in grado di propagare la mutua informazione tra gli elementi del sistema attraverso ampie regioni, riuscendo ancora a limitare le disinformazioni (danni) in spazi ristretti-, ma sufficientemente caotico da mostrare i volumi che confinano comportamenti differenti non più nettamente distinti, ma sinuosi e intersecantisi nello spazio dei parametri. In questi casi, piccoli cambiamenti nei parametri di funzionamento, o modeste perturbazioni possono condurre a biforcazioni nel comportamento del sistema tali da sottendere successivi improvvisi e imprevedibili cambiamenti radicali di traiettorie e attrattori. In questa situazione si riduce il grado di modularità strutturale e/o funzionale con sottosistemi sempre più interconnessi e interagenti. Manifesta, quindi una minore stabilità omeostatica, assumendo comportamenti, in presenza di perturbazioni, che si muovono in volumi, nello spazio delle sue possibilità, non ben confinati, e laddove riesca a ritornare allo stato iniziale mostra un equilibrio stazionario instabile. Nel linguaggio dei sistemi dinamici non ha un esatto analogo, potendosi solo riferire in termini di complessità propriamente detta; volendo comunque darne un riferimento il comportamento in questa fase corrisponde agli attrattori periodici instabili a crescente dimensionalità. Nel campo delle classi di universalità degli automi cellulari questa fase corrisponde alla classe IV, e, in ambito fisico, alle transizioni di fase di secondo ordine solido/fluido.
La complessità caotica è la fase in cui i comportamenti complessi controllabili sembrano essere preclusi. Il sistema è strutturalmente instabile: anche partendo da condizioni iniziali deterministiche, piccoli cambiamenti, nella struttura e/o nella funzione, causano quasi sempre vasti e sproporzionati cambiamenti nel comportamento dell’insieme, di cui non si riesce più a rintracciare caratteristiche modulari, con sottosistemi così fortemente interagenti da annullarne, tendenzialmente, le individualità. Minime alterazioni delle condizioni iniziali danno luogo a drastici mutamenti nella risposta, in cui può essere esibito ogni comportamento, anche il più imprevedibile, e la mutua informazione utile tra le variabili del sistema decresce fino a 0. In queste condizioni la tendenza a riassumere il comportamento iniziale è sostanzialmente persa, ed ogni perturbazione tende a diffondersi in spazi vasti e in tempi lunghi. Nel linguaggio dei sistemi dinamici i comportamenti in questa fase corrispondono agli attrattori "strani"8;(caos basso-dimensionale) (30.Ruelle, 1979) e agli attrattori alto-dimensionali. Corrisponde analogicamente anche agli automi di classe III e al regime fluido.
Nota
8 Individuato dal metereologo Lorenz nel 1963 (anche per questo conosciuto come "attrattore di Lorenz") e definito da Ruelle (1979), č un attrattore di dimensione frattale,- non intera cioč-, su cui il moto di un punto presenta il fenomeno della "dipendenza sensibile dalle condizioni iniziali", -netta divergenza di traiettorie di due punti arbitrariamente vicini-, e che benché di dimensione finita puň descrivere un numero infinito di modi (oscillazioni), cioč totalmente imprevedibili.
Va sottolineato che i 3 gradi di complessità descritti non inquadrano staticamente volumi confinati di comportamento, all’interno di cui, una volta organizzato il suo livello strutturale e funzionale, un sistema dinamico complesso dissipativo rimanga indeterminatamente nel tempo. Individuano, piuttosto, ambiti statisticamente differenti, che denotano la comparsa o scomparsa di proprietà nuove o comunque differenti, di comportamenti variamente imprevedibili e finalisticamente utili, attraverso cui uno stesso sistema può guidare, in particolari condizioni, le sue transizioni strutturali e/o funzionali. Proseguendo l’analogia già presentata con gli stati della materia ordinaria, così come altri autori hanno fatto anche se per altre categorie (20.Langton, 1986; 21.Langton, 1990; 5.Crutchfield and Young, 1990; 17.Kauffman, 1993) e purché non sia presa troppo letteralmente, possiamo riconoscere alla fase di complessità subcritica l’affinità con lo stato solido (semplice, ordinato, stabile), alla fase di complessità caotica lo stato fluido (imprevedibile, disordinato, instabile) e a quella di complessità critica la fase di transizione solido/fluido (complessa ma sufficientemente ordinata, variamente imprevedibile ma ancora sufficientemente stabile). E’ plausibile che un sistema biologico possa raggiungere i suoi optima, sia in termini di capacità di assumere i comportamenti e assolvere gli obiettivi più complessi, sia come capacità di evolvere più adeguatamente in un mondo che cambia, in un regime ordinato ma vicino al margine del caos.
È sempre stata opinione prevalente che la formazione, lo sviluppo, l’organizzazione e l’evoluzione del sistema nervoso sia frutto soltanto di una finissima programmazione genetica in cui i singoli neuroni, come componenti di un puzzle pre-programmato, sono semplici tasselli di un disegno deterministicamente pre-definito. Come tale, lo studio delle "finestre di apprendimento", delle modalità cioè con cui l’impianto neurale organizza dinamicamente il suo comportamento nei confronti del mondo esterno, sembra appannaggio, prevalentemente, dell’ambito psicologico. Il rilievo di una capacità di sviluppo e organizzazione autonoma rispetto alla programmazione genetica, con soluzioni e modelli di funzionalità originali, fa emergere l’evidenza di, e l’interesse per la multiforme complessità insita nella struttura e nella funzione dei sistemi viventi, complessità riscontrabile su qualsiasi scala li si consideri. E con essa la necessità di un approccio interdisciplinare, che sappia coinvolgere le scienze esatte, quali matematica, fisica, informatica, nello studio dei fenomeni biologici.
Se la riflessione si rivolge, poi, al campo algologico, la richiesta si fa urgente. Il fenomeno "dolore", infatti, nonostante dal 1965 si sia aperta la strada ad una serie di ricerche che hanno portato alla luce un numero sempre crescente di sistemi interni di modulazione dell’input sensoriale dolorifico, continua ad essere compreso solamente nei termini di funzione sensoriale lineare. I conseguenti, e innegabili, progressi terapeutici hanno beneficato soprattutto l’espressione acuta del sintomo, lasciando estesi settori del dolore cronico in preda ad enigmatici, non confortanti, risultati.
In questo ambito, il metodo analitico si è sempre mosso al di fuori di un qualunque modello descrittivo e predittivo di riferimento, se non parziale.
È sicuramente da sottolineare che nessuna formula matematica può simulare esattamente il comportamento di un sistema vivente, e che qualsiasi modello è solo un’approssimazione che facilita nella descrizione e nella comprensione della realtà, senza mai, però, poterne esaurire la variabilità e complessità. Pur nei limiti già osservati, la modellizzazione che delineeremo ci consente di circoscrivere un’area in cui la fenomenologia del dolore riceve caratterizzazioni inaspettate, anche se per il momento solo analogiche e qualitative. I rilievi che ne derivano, infatti, hanno il valore di affermazioni, certo di carattere generale, in grado comunque di ricomprendere l’espressione dolorosa con chiavi interpretative adeguate alla complessità del fenomeno.
Il sistema Nd/d è un sistema strutturato ad architettura gerarchica, composto da 3 sottosistemi: sensoriale-discriminativo, motivazionale-affettivo e cognitivo-valutativo (23.Melzack and Casey, 1968). L’evento doloroso, cioè, determina, nel soggetto in cui si verifica, un’esperienza complessa e diversificata che include:
In condizioni basali e a fisiologia integra, tutto il sistema è organizzato ad alto grado di modularizzazione, con una attività che privilegia, specie nelle risposte a bassa frequenza, le interazioni intra- piuttosto che quelle inter-modulari.
Il suo comportamento dinamico è sottoposto a processi di regolazione secondo un modello omeostatico, che essenzialmente può essere descritto:
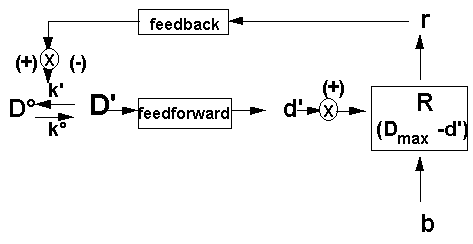
Esso è fondamentalmente un sistema di regolazione, a circuiti multipli, a controllo in feedback/feedforward, non lineare: esiste la variabile considerata (in questo caso la variabile Do-D'), che può modificarsi in modo reversibile, grazie all’esistenza di meccanismi che spingono la reazione o la trasformazione nelle due direzioni (nel caso considerato, Do--->D' e D'--->Do). Chiameremo questi meccanismi "sistemi effettori", essendo essi capaci di effettuare la trasformazione o la modificazione del parametro considerato. Lo stato di D e D' dipendono dal relativo disequilibrio dell’attività dei due sistemi effettori opposti (denominate rispettivamente k e k'). Il sistema omeostatico è qui rappresentato da un anello di retroazione (feed-back) in cui l’informazione sul risultato di una trasformazione o di una oscillazione di attività viene rimandata, riveduta e corretta, all’ingresso del ciclo. Non esistono solo anelli di retroazione negativi (che garantiscono la stabilità), ma anche anelli positivi, di autoamplificazione, in cui il prodotto della reazione contribuisce ad accelerare la trasformazione. L’elaborazione umana del dolore è, inoltre, rafforzata dal controllo in feedforward ("informazione anticipata" - nel nostro caso uno di tali circuiti è dato dal "central control trigger", sistema specializzato di fibre a grosso diametro e rapida conduzione, formato dalle vie di proiezione delle colonne dorsali e dal sistema post-sinaptico delle colonne dorsali), che prevede le possibili conseguenze del dolore e delle esperienze dolorose, intervenendo a pianificare preventivamente scenari alternativi. L’apprendimento, infatti, come componente primaria del dolore spiega anche l’individualità dei suoi effetti, a parità di intensità fisica, su persone diverse.
Affinché la normale omeostasi venga garantita, l’attività dei sistemi effettori non avviene senza controlli reciproci e senza coordinamento con altri sistemi: a questo scopo, in ogni sistema omeostatico sono inseriti uno o più sistemi regolatori (R), che svolgono il ruolo fondamentale di determinare quale disequilibrio Do/D' debba essere raggiunto o mantenuto in ogni momento, integrando lo stato di Do/D' con le esigenze del resto dell’organismo. Per far questo, il sistema regolatore deve "monitorare" lo stato di Do e D', ricevendo da essi adeguata segnalazione (d'), e deve produrre un segnale di ritorno (r) che viene recepito in senso stimolatorio o inibitorio dai sistemi effettori.
Il sistema regolatore è quindi capace di controllare i sistemi effettori ed è collegato ad altri sistemi da cui riceve informazioni utili alla scelta del controllo da effettuare (b).
Associato al sistema regolatore vi è anche un controllo intrinseco, che potrebbe dirsi la memoria dei valori normali di Do e D'. I valori "normali" sono i limiti massimo e minimo della variabile Do, al di sopra ed al di sotto dei quali il sistema regolatore si mobilita emettendo un segnale che influenza le attività dei sistemi effettori, nel senso di riportare i valori nella norma. Nello schema si è espresso tale concetto con la formula (Dmax-d'), intendendo con questo il fatto che il sistema regolatore si attiva in base al confronto tra il valore massimo di Do tollerabile (Dmax) ed il segnale che indica la situazione attuale o potenziale (d'). In questo schema, tanto più alto sarà d' rispetto ad Dmax, tanto più intensa sarà la produzione di r e quindi l’inibizione del sistema effettore Do--->D'. I limiti superiore ed inferiore cui si fa qui riferimento sono dati dalle condizioni strutturali intrinseche del sistema regolatore, non sono per tutti i sistemi invariabili, anzi molti sistemi omeostatici si adattano su diverse soglie di risposta quando compaiono malattie croniche, o di fronte a stimoli ripetitivi. I sistemi regolatori sono quindi "plastici": a questo livello si verificano vari eventi regolativi come i fenomeni di ipersensibilità (ad es.: aumento di recettori, "priming") o di adattamento (ad es.: "down-regulation", inibizione da contatto, etc.).
Il sistema Nd/d appartiene, insieme a molti altri sistemi tra cui quello immunitario, ad una vasta, interdisciplinare classe di sistemi complessi che i ricercatori dell’Istituto di Santa Fe (12.Holland, 1992; 13.Holland, 1995; 9.Gell-Mann, 1994; 31.Singer and Morowitz, 1995) hanno raggruppato sotto il nome di sistemi adattativi complessi (CAS -complex adaptive systems). Tali sistemi, pur nella loro marcata diversità, esibiscono un insieme di significative caratteristiche comuni da rendere possibile e probabile che principi generali comuni spieghino le loro dinamiche.
Un CAS raccoglie informazioni sull’ambiente che lo circonda, su se stesso e sui suoi comportamenti, in modo che questa attività di monitoraggio, pur se apparentemente stazionaria, in effetti è mutevole nel tempo secondo modelli che portano il sistema ed il suo ambiente ad essere coevolventi. Esso è specificato dalle seguenti caratteristiche generali:
È importante distinguere ciò che è adattativo da ciò che è il risultato di un processo di adattamento, perché quest’ultimo può essere spesso maladattativo.
Se osserviamo, infatti, il comportamento del sistema Nd/d è facile notare come il più costante modello di maladattamento è il prevalere dell’efficienza di segnalazione sull’efficacia di risposta. Vale a dire si realizza uno schema in cui la ben realizzata segnalazione d’allarme per la sopravvivenza dell’individuo è più importante della sopravvivenza stessa. Il meccanismo che riteniamo il più delle volte sotteso è quello che abbiamo definito mutazioni di connettività intra- e inter-neuronali, intra- e inter-modulari, a cui occorre aggiungere quello delle mutazioni funzionali: instabilità dei loops omeostatici per disfunzione (in iper o in ipo) dei processi di feedback positivi e/o negativi (ad es. iperattività della funzione facilitatoria del sistema simpatico o degli inputs facilitatori reticolari; alterazione dei meccanismi inibitori; processi di trasformazione neuronale quale quelli che portano un neurone nocicettivo specifico a trasformarsi in WDR). Altra caratteristica che abbiamo rilevato essere frequentemente implicata nella selezione di uno schema maladattativo è una condizione di risposta ad alta frequenza (stimoli particolarmente intensi, o particolarmente prolungati nel tempo, o ad alta reattività intermodulare), in cui, tra l’altro, il meccanismo più sensibile sembra essere quello di memorizzazione e comparazione degli impulsi. Il risultato che spesso si produce nell’abbinamento di più meccanismi maladattativi è quello di circuiti riverberanti, più o meno locali, ad attività automantenentesi, afinalistici rispetto alle funzioni biologiche del sistema.
Il dolore acuto e cronico differiscono nella loro eziologia, fisiopatologia, funzione biologica, diagnosi e terapia. E non sembra inutile sottolineare che il dolore cronico non è un dolore acuto che persiste.
La cornice concettuale sin qui esposta ci consente di inquadrare nuove ipotesi interpretative (modelli) dell’espressione dolorosa. I rilievi che ne derivano, infatti, hanno il valore di affermazioni, certo di carattere generale, in grado comunque di ricomprendere l’espressione dolorosa con chiavi interpretative adeguate alla complessità del fenomeno.
1. Il sistema nondolore-dolore è un sistema adattativo complesso, organizzato secondo circuiti omeostatici non-lineari, non-ergodico, in cui quindi l’elaborazione dell’informazione biologica avviene tanto in parallelo asincrono, quanto, almeno a livello percettivo corticale, in parallelo in priorità d’attenzione.
2. In fase di complessità subcritica il sistema Nd/d, strutturalmente e funzionalmente integro, è organizzato omeostaticamente in sottosistemi funzionalmente isolati con controllo prevalente in feedback negativo. Il sistema confina spontaneamente i suoi comportamenti dinamici in un volume relativamente piccolo all’interno del suo spazio delle fasi, realizzando schemi che abbinano efficienza di segnalazione d’allarme con efficacia di risposta adattativa. La progressione temporale dei suoi modelli di attività segue attrattori punto o periodici stabili: l’informazione (nondolore/dolore) percorre cioè dei cicli finalizzati ricorrenti, tendendo fortemente a ritornare allo stesso comportamento iniziale e a mantenerlo una volta finita la perturbazione: è quindi omeostaticamente stabile. In tale fase il sistema si comporta come lineare e vi può effettivamente essere un equilibrio statico. In questa ottica la comparsa di un dolore acuto non è altro che lo spostamento a destra del parametro oscillante nondolore-dolore, cui il sistema omeostatico contrappone un feedback di controllo, che, in presenza di una perturbazione transitoria, riarmonizza il sistema.
3. Il dolore cronico esprime sempre un modello di maladattamento, sotto la spinta di mutazioni di connettività e funzionali, e spesso in condizioni di risposte ad alta frequenza; ed il passaggio ad un grado di complessità superiore, tipicamente alla fase di complessità critica. In questa situazione si riduce il grado di modularità strutturale e/o funzionale con sottosistemi sempre più interconnessi e interagenti. Manifesta, quindi una minore stabilità omeostatica, assumendo schemi non adattativi ma di (mal)adattamento, in presenza di perturbazioni in grado di modificare, attraverso meccanismi di plasticità, aspetti strutturali e/o funzionali dei circuiti neuronali: sintesi di nuove proteine recettore e di canale, formazione di nuove connessioni ecc., che ne modificano la loro meccano- e chemosensibilità, e la loro capacità organizzativa. In tutto questo, un ruolo centrale sembra essere assunto dal ganglio della radice dorsale (DRG), almeno nei quadri di dolore neuropatico (10.Gupta and Raja, 1996), sede di produzione di impulsi ectopici, di proteine recettore e di canale, target di stimolazione simpatica attraverso le nuove connessioni che dagli assoni noradrenergici perivascolari raggiungono il ganglio stesso. Si realizza, così, un modello di comportamento a circuiti riverberanti, automantenuti da schemi che il sistema realizza per adattarsi alla nuova situazione. Nel linguaggio dei sistemi dinamici, questa situazione non ha un esatto analogo, potendosi solo riferire in termini di complessità propriamente detta; volendo comunque darne un riferimento il comportamento in questa fase segue attrattori periodici instabili a crescente dimensionalità. Il passaggio ad una complessità superiore può interessare parte (che è il caso più frequente), una o più reti (sottosistemi), riducendo sempre più la modularità del sistema, con attrattori che occupano volumi sempre più grandi per l’innesco di meccanismi autoamplificatori, fino a poter coinvolgere sistemi sensoriali adiacenti ( disestesie, allodinia, ecc.).
4. Se il dolore cronico è espressione di una patologia da maladattamento, la strategia terapeutica non può che prevedere, come momento fondamentale, l’intervento sui meccanismi di plasticità centrale, che sembrano essere il pivot di ogni modifica strutturale/funzionale, oltreché dei meccanismi di apprendimento e di realizzazione degli schemi adattativi o maladattativi. Al momento, sembra che ogni intervento terapeutico non sia in grado di proporre un’azione "rimodellatrice", ma solo inibitoria. Inibizione che pur se in grado, talvolta, di dare tregua al dolore, viene spesso bypassata dalle capacità riorganizzative del sistema. Implicazioni che non crediamo possano essere ristrette unicamente al trattamento del dolore neuropatico. 4 sembrano, a nostro avviso, i presupposti di fallimento, nel lungo termine, di una terapia (specie farmacologica):
Introdurre il concetto di complessità nel campo della biologia e della medicina, significa indubbiamente intraprendere un viaggio conoscitivo e descrittivo in un paesaggio dove gli spazi simbolici (punti di vista, livelli di lettura) in cui la realtà si esprime possono assumere un aspetto inconsueto rispetto alle più praticate metodologie analitiche.
Nella critica ai dogmi riduzionisti abbiamo opposto sottosistemi che si muovono in uno spazio ideale (dinamica non-lineare), a singole particelle elementari che si muovono in uno spazio reale (meccanicismo), constatando che nel caso di sistemi complessi occorrono leggi efficaci diverse dalle leggi microscopiche per darne una descrizione del comportamento macroscopico.
Il sistema Nd/d ne rappresenta sicuramente un’evidenza. Interpretato sempre come un fenomeno lineare, mostra invece, nella modellizzazione di complessità, la capacità di comprimere le regolarità individuate in uno schema, in un modello interno, cioè, di comportamento e di comprensione della realtà, che è tutt’altro che lineare.
Garante, insieme al sistema immunitario, della nostra identità, si rivela essere un sistema adattativo complesso che raccoglie informazioni sull’ambiente che lo circonda, su se stesso e sui suoi comportamenti, in modo che questa attività di monitoraggio, pur se apparentemente stazionaria, in effetti è mutevole nel tempo secondo modelli che portano il sistema ed il suo ambiente ad essere coevolventi.
Il dolore acuto e quello cronico risultano così espressioni radicalmente diverse: lineare, fisiologico, finalizzato il primo; non-lineare, maladattativo, con l’unico fine probabilmente di circoscrivere, finché riesce, le disinformazioni il secondo.
Richiamandoci a quanto scritto altrove, se ci rifacciamo ad uno schema [R (Realtà) - M (Apparato di misura) - S (Simboli)] (28.Popper, 1969) non potremmo che concludere che, essendo noi schermati da M, ed essendo questo sempre provvisorio in quanto legato ad una specifica teoria, tutto quello che raccogliamo in S è doxa (opinione) e non epistème (scienza vera), come invece riteneva Aristotele. Che M sia inadeguato per descrivere nuove situazioni, per noi vuol solo dire che la verità di M è parziale, e va integrata con altre. Occorre allora organizzarsi a livelli diversi d’indagine, integrandone le informazione.
 ©Copyright Progetto Salute
©Copyright Progetto Salute